慶應発サイエンス
閉経後骨粗鬆症を発症させる因子の同定に成功
閉経後骨粗鬆症とは
骨粗鬆症とは、文字通り骨がすかすかになって脆くなり、折れやすくなっている状態のことです。閉経や加齢、ステロイドなどの薬剤投与、甲状腺機能亢進症などの疾患など、実に様々な要因により、骨粗鬆症は発症します。骨の量や強度は、X線を使って計測する骨密度で類推することができ、この骨密度が若い人の平均値の70%以下、つまり30%以上低下すると骨粗鬆症と診断されます。骨密度の計測値から推測する骨粗鬆症の患者数は、国内で現在約1300万人と考えられ、さらに増加し続けています。
骨粗鬆症による骨折の発生
骨粗鬆症で骨が脆くなっているだけでは、症状がでることはほとんどありません。問題は、骨が脆いので骨折しやすくなっていることで、骨折して初めて骨粗鬆症に罹患していることに気がつくことも稀ではありません。骨粗鬆症による骨折で最も深刻な骨折は大腿骨近位部骨折といって、太ももの骨の付け根で折れる骨折です(図1)。この大きな骨が折れると、当然立ったり歩いたりできないので、救急車で病院に搬送されることになります。ギプスで固定するような治療では、長くベッド上安静になるため、高齢者では認知症や褥瘡の発症など、さらに深刻な問題が出てきてしまうので、麻酔をかけて手術で金具を使って固定し、早く立てるようにすることも少なくありません(図1)。ただ、高齢者への手術はリスクを伴います。もともと高齢ということもありますが、手術後1年以内に亡くなる方が約30%、5年以内では50%以上の方が亡くなる、とされています。骨粗鬆症があると、ちょっとした転倒で、このように深刻な大腿骨近位部骨折につがるので、骨粗鬆症の理解と治療が骨折予防にはとても大切になります。
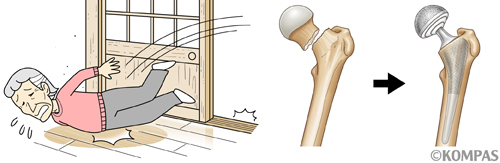
図1.高齢の骨粗鬆症患者では、ちょっとした段差につまずいて転倒し(左)、太ももの付け根の骨折を起こしやすくなります(中)。骨折を起こすと、高齢者でも手術により金属等を使って治療するケースが多くなります。
骨粗鬆症の発症原因
骨粗鬆症の最も大きな要因の1つは閉経であり、また加齢です(図2)。70歳代以降の女性の二人に一人が骨粗鬆症に罹患していると報告されています。閉経は、50歳頃に月経が終了することですが、閉経に伴い急速に骨密度が低下することが知られています。これは、女性ホルモンであるエストロゲンが閉経により急速に失われ、それに伴い、骨を吸収する破骨細胞の活性が強まり、骨密度の低下が起こることによります。閉経後の骨密度低下はエストロゲンが減少することが病態であるので、エストロゲンそのものを閉経女性に投与すると、閉経前のように骨密度が保てるようになります。しかし、エストロゲンそのものの投与は子宮体癌や乳がん、血栓症のリスクを増大させるため、現実には実施することができません。そのため、現在の治療の多くは、閉経により強くなった破骨細胞の活性を抑制する薬を投与することであり、一部、逆に相対的に弱くなった骨を作る側の細胞である骨芽細胞を活性化させる薬を患者さんに投与することです。
閉経前では破骨細胞による骨吸収と骨芽細胞による骨形成のバランスが一致し、骨量は一定に保たれますが、閉経後のエストロゲン欠乏状態では破骨細胞の活性が骨芽細胞の活性より相対的に強まり骨量が減少するようになります(図3)。しかし、なぜ閉経によって破骨細胞が活性化するのか、そのメカニズムは知られておらず、破骨細胞の活性化の理由がわからずに治療しているのが現状でした。この結果、薬の投与による有害事象も発生することがありました。
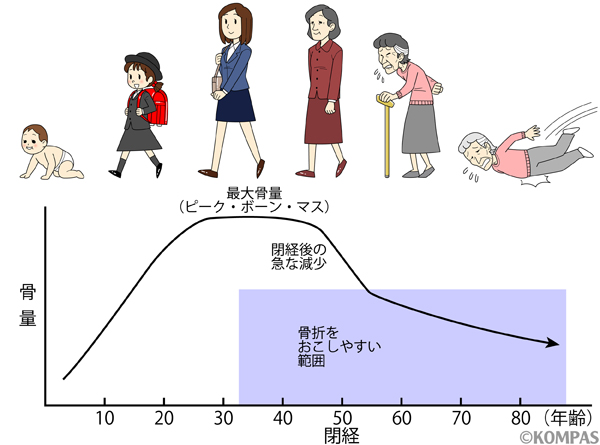
図2.女性は、50歳前後に起こる閉経により急速に骨量が減少し、その後加齢によりややなだらかに骨量が減少する、という2段階の骨量減少様式を示します。つまり、女性では閉経と加齢が骨粗鬆症の大きな要因と言えます。
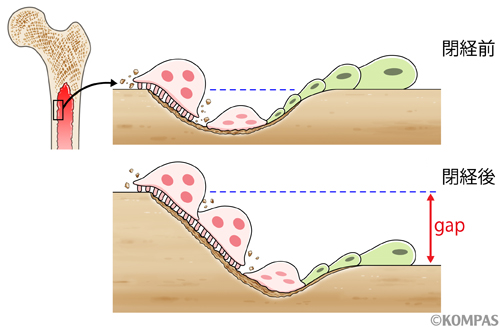
図3.閉経前は、骨を吸収する破骨細胞と、骨を形成する骨芽細胞との、絶妙なバランスにより骨量は常に一定に保たれています(上段)。しかし、閉経によるエストロゲン欠乏により、破骨細胞の活性が骨芽細胞に対して相対的に優位になることで、骨量が減少するようになります(下段)。
これまでの破骨細胞研究
閉経後の骨粗鬆症発生に中心的な役割を担う破骨細胞のことを知るために、破骨細胞に発現する遺伝子や蛋白を網羅的に調べる研究がたくさん報告されています。マウスの骨髄から採取した破骨細胞前駆細胞を培養して大量に破骨細胞を作り、遺伝子や蛋白の発現を解析し、破骨細胞に特徴的な分子を特定します。我々もそのような研究手法で、いくつかの重要な遺伝子を同定してきました。しかし、この手法では、もともとあるエストロゲンがなくなる、ということを再現させることが難しいという難点がありました。また、破骨細胞は骨の表面にのみ存在しますが、骨の表面は極めて酸素濃度が低い低酸素環境であることが知られていますが、これまでの試験管での破骨細胞の形成系では、低酸素、というキーワードも欠けていました。
破骨細胞に発現するHIF1αが閉経後骨粗鬆症の発症を誘導する
そこで、破骨細胞に発現する低酸素応答性因子を検索したところ、低酸素応答性転写因子であるHypoxia inducible factor 1 alpha (HIF1α)が発現することを見出しました。HIF1αは遺伝子で発現しても、酸素があると蛋白レベルでは分解されてしまいます。試験管での培養は正常酸素濃度で行うため、HIF1α蛋白は破骨細胞では検出されませんが、低酸素培養装置で試験管の培養を行うと、破骨細胞にもHIF1α蛋白が検出されるようになります。また、大変興味深いことに、閉経前のマウスの骨を取り出して、免疫染色という手法でHIF1α蛋白の発現をみてみても、破骨細胞にはほとんど検出されませんが、マウスの両側の卵巣を摘出して人工的にエストロゲン欠乏の閉経状態にすると、骨密度の低下とともに骨の中の破骨細胞にHIF1α蛋白が検出されるようになりました(図4)。つまり、エストロゲンも酸素と同じようにHIF1α蛋白を制御していることが示唆されました。
つぎに、エストロゲン欠乏によって破骨細胞の中に検出されるようになるHIF1α蛋白が、破骨細胞の活性化から骨密度低下の要因となっているかを調べるために、破骨細胞にだけHIF1αが発現しなくなる特殊な操作をしたマウスを用いて、卵巣摘出によるエストロゲン欠乏状態にしてみると、骨密度の低下がかなり抑えられることがわかりました。さらに、HIF1α阻害剤を卵巣摘出によるエストロゲン欠乏状態のマウスに投与すると、破骨細胞の活性化や骨密度の低下が完全に抑えることができました(図4)。
破骨細胞は、通常の骨のリモデリングと呼ばれる新陳代謝に重要な役割を担っており、この活性をもまるごと抑制する骨粗鬆症治療用の薬剤の投与では、骨のターンオーバーが起こらない状態になってしまいます。しかし、HIF1α阻害剤は、エストロゲン欠乏により異常に活性化した部分だけを標的にするので、生理的な骨のターンオーバーは保たれます。我々はHIF1αという新たな閉経後骨粗鬆症の治療標的と、その治療剤を手に入れたのです。
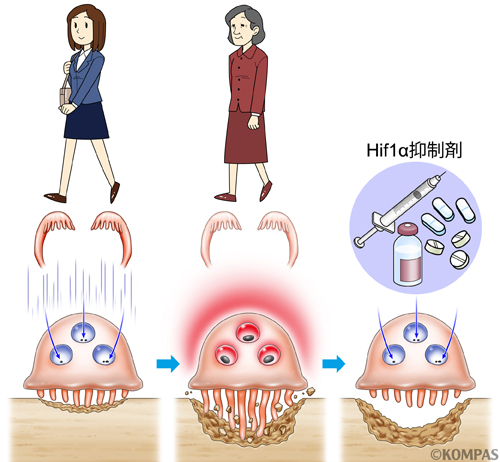
図4.破骨細胞は体の中でも特に酸素が薄いところに存在し、低酸素応答性のHIF1αを発現してしまいますが、閉経前はHIF1αはエストロゲンにより常に抑制されています(左)。しかし、閉経によりエストロゲンが欠乏すると、この抑制が効かなくなり、HIF1αの活性化により破骨細胞の活性が上昇し、骨量が減少するようになります(中)。HIF1α阻害剤の投与により、エストロゲンの代わりにHIF1αを抑制すると、閉経により異常に活性化した部分のみを抑制し、破骨細胞の状態を閉経前と同じ状態にすることができます(右)。この際、生理的な骨のターンオーバーに必要なベースの破骨細胞の活性は、閉経前のように保たれます。
今後の展望
現在、骨折のない閉経後骨粗鬆症の治療は、骨密度の若年平均値から30%以上低下して初めて治療が開始できますが、今日の治療剤では3年間の治療でだいたい腰椎で良くて7~10%、大腿骨だと3~5%程度骨密度があがる程度です。しかし、破骨細胞でHIF1αが活性化してきたことを検知できれば、骨密度が大きく低下する前にHIF1α阻害剤で治療ができ、骨密度を若い時のまま維持することもできるようになるかもしれません(図5)。
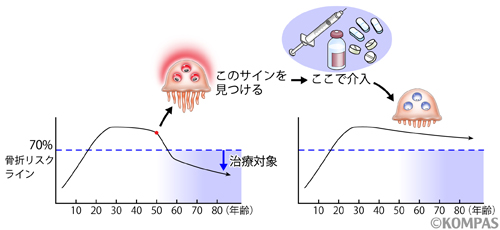
図5.閉経後、骨量が大きく低下してしまう前に、破骨細胞が活性化した状態を検出することができれば、この時点でHIF1α阻害剤による治療を開始することで、閉経による大きな骨量減少を未然に防ぐことができるようになるかもしれません。
参考文献
HIF1α is required for osteoclast activation by estrogen deficiency in postmenopausal osteoporosis.
Miyauchi Y, Sato Y, Kobayashi T, Yoshida S, Mori T, Kanagawa H, Katsuyama E, Fujie A, Hao W, Miyamoto K, Tando T, Morioka H, Matsumoto M, Chambon P, Johnson RS, Kato S, Toyama Y, Miyamoto T.
Proc Natl Acad Sci U S A. 2013 Oct 8;110(41):16568-73. doi: 10.1073/pnas.1308755110. Epub 2013 Sep 10.
http://www.pnas.org/content/110/41/16568.long![]()


SHARE