慶應発サイエンス
脳梗塞を悪化させる炎症の仕組みを解明
脳梗塞と炎症
脳梗塞とは、脳の血管が詰まるなどして脳血流が乏しくなること(虚血)によって脳組織が壊死に陥る病態です。日本では継続的に治療を受けている患者総数120万人と言われ、がんに匹敵する患者数です(「平成20年患者調査」厚生労働省)。脳卒中(脳梗塞を含む)は死亡率が高い(年間13万人)だけでなく、助かっても寝たきりになるなど患者の生活の質(QOL)が非常に悪いことが特徴です。これまでの脳梗塞の治療は発症直後(4~5時間以内)に血栓溶解薬を投与するなど、限られた治療法しかありませんでした。しかし梗塞後徐々に炎症が起こって脳組織が浮腫を起こして腫れることにより、梗塞巣が広がったり、病状が悪化したりする事があり、これらを抑制することも重要と考えられています。
脳梗塞は脳の血流障害(虚血)による脳組織の損傷ですが、脳組織の壊死によって発症早期に大きな炎症がひき起こされます。炎症はそれから数日間続き、脳梗塞の病態の悪化に関連している可能性が考えられます。まず梗塞直後に炎症をひきおこす細胞としてマクロファージが侵入し、次にやや遅れて3日目をピークにリンパ球が侵入します。私たちはまず脳梗塞の病態におけるリンパ球の役割を明らかにし、さらに梗塞直後のマクロファージの活性化のメカニズムを明らかにしました。これらの作用を調節することにより脳神経の保護に結び付け、新たな治療法を開発できる可能性があります(図1)。
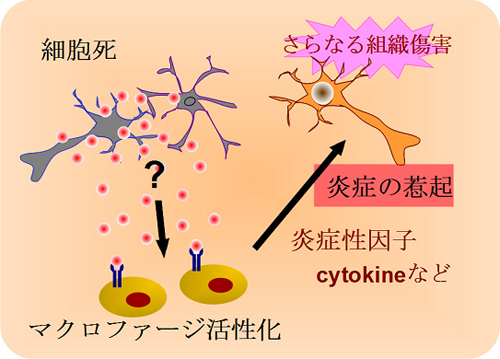
図1 脳虚血による炎症惹起のメカニズム
脳虚血によって組織の細胞死が起こると、何らかのメカニズムによって、脳内に浸潤した血液細胞が活性化され、炎症性因子サイトカインを産生することによりさらなる組織傷害が惹き起こされる。
マクロファージ活性化のメカニズム解明から新しい治療法へ
脳梗塞を悪化させる炎症性サイトカインとその産生細胞
炎症はマクロファージやリンパ球が産生するサイトカインによって拡大し組織に傷害を与えます。多数のサイトカインのなかで、どのサイトカインが重要なのかを明らかにするために、IL-6(インターロイキン‐6), IFNγ(インターフェロンγ)、IL-17,IL-23などのサイトカイン遺伝子を破壊した遺伝子欠損マウスを使ってマウスの脳梗塞モデルを作製し、観察しました。その結果IL-23とIL-17を欠損した場合、脳梗塞体積の減少が認められました(図2)。
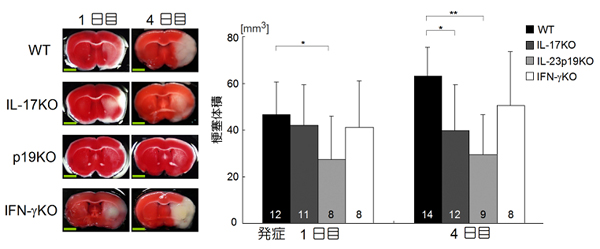
図2 IL-17、IL-23遺伝子欠損マウスは脳虚血による組織障害が減弱する
実験的に脳梗塞を作成したマウスを用いてIL-17、IL-23、IFN-γの作用を検討した。脳梗塞巣は白い部分で示される。IL-17遺伝子欠損(KO)マウスでは発症4日目に、IL-23p19遺伝子欠損(KO)マウスでは発症1日目から、無処置の野生型(WT)マウスよりも著明な脳梗塞体積の減少を認めた。
次にどの細胞がサイトカインを産生するのかを調べたところ、IL-23は発症1日目に脳梗塞巣内に浸潤したマクロファージによって産生されていました。さらに、このIL-23によって、発症3日目に同部に浸潤したT細胞からIL-17の産生が誘導されていました。T細胞にはいくつか種類がありますが、これまでIL-17の主な産生細胞はTh17(ヘルパーT細胞)と考えられて来ました。ところが、驚くべきことに、このモデルではIL-17の主な分泌細胞は当初想定していたTh17ではなく、感染防御においてはTh17よりも比較的早期に活躍するγδT細胞でした(図3)。γδT細胞もIL-23の刺激によってIL-17の産生を開始することが知られています。すなわち
脳梗塞 → マクロファージ → IL-23 → γδT細胞 → IL-17 → 炎症と傷害の拡大
と梗塞巣から炎症が拡大していく機序が明らかとなりました(文献1)。
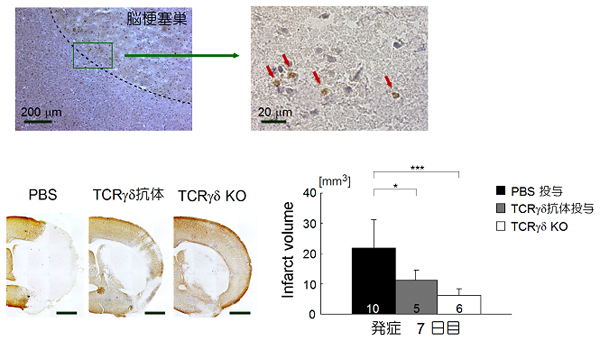
図3 脳梗塞巣におけるγδT細胞の浸潤と組織障害
上: 脳梗塞巣内におけるγδT細胞の存在を免疫染色法によって検出しました。γδT細胞は梗塞周辺部に局在していました。
下: 野生型マウスにPBSを投与したマウス、γδT細胞を除去する抗体を投与したマウスおよびTCRγδ遺伝子欠損(KO)マウスをそれぞれ用いて実験的に脳梗塞を作成した。脳梗塞巣は白い部分で示されています(MAP-2免疫染色)。γδT細胞の除去によって梗塞体積の著明な減少を認めた。このことから炎症の拡大にδT細胞が関与していることが示唆されました。
γδT細胞を標的とした脳梗塞治療戦略
免疫染色法によって染まるγδT細胞は、脳虚血による神経細胞死が進行している梗塞巣の周辺部に局在していることを証明しました。これらのγδT細胞を抗体によって除去したマウスでは、何も前処置をしない野生型マウスよりも脳梗塞体積が小さくなることが明らかとなり、さらに脳梗塞発症24時間が経過してからであっても、抗体投与によってγδT細胞を除去することにより脳梗塞体積の減少が見られました。またIL-23中和抗体を投与することによっても脳梗塞体積を減少させることができました(文献2)。
さらに、私たちは脳梗塞巣内へのT細胞の浸潤を阻止することにより脳梗塞体積を減少させることにも成功しました。FTY720(Fingolimod)はT細胞のリンパ組織からの遊走、炎症組織への浸潤を阻害する薬剤として知られています。現在、多発性硬化症に対して保険適用が認可されています。FTY720を静脈内投与すると脳梗塞巣内に浸潤したγδT細胞の数が減少し、脳梗塞体積は著明に減少しました。この結果はγδT細胞が脳内に侵入することが梗塞の拡大に重要な因子であるという仮説を支持するものであり、またFTY720が脳梗塞の増悪を防ぐ薬剤として使用できる可能性を示唆しています。
脳組織中で炎症を起こすPrxファミリーたんぱく質
脳梗塞におけるマクロファージTLRの意義
ではIL-23はどのようにしてマクロファージから産生されるのか? それが次の疑問でした。マクロファージは主にToll-like receptor (TLR)という受容体を使って細菌を認識しており、TLRの発見は2011年のノーベル賞の対象となりました。TLRには10種類以上がありますが、私たちは遺伝子破壊マウス(ノックアウトマウス)を用いてこのうちTLR4やTLR2が脳梗塞後の炎症に関与していることを示しました。またTLR2欠損とTLR4欠損を比べるとTLR2欠損のほうが効果がより大きく、さらにTLR2/4両欠損マウスでは梗塞体積の減少がさらに大きくなり相加効果が見られました。しかし脳は本来無菌的で清潔な臓器であり、細菌やウイルスなどの外敵は通常存在しません。したがって、TLRに認識される因子が脳組織の中にもともと存在しており、壊死に陥った時にはその因子が細胞外にばらまかれて、TLRを介してマクロファージを刺激して活性化させると考えられます。このような因子は、ダメージ関連分子パターン(damage-associated molecular patterns: DAMPs)と呼ばれています。これまで脳内のDAMPsとしてはhigh mobility group box 1 (HMGB1)、heat shock protein、β-amyloidなどが研究されてきましたが、炎症の引き金となる重要なDAMPsは不明でした。
脳梗塞においてTLRを活性化するPrx
私たちは脳組織中に存在する新規のDAMPsの発見をめざしました。まず、脳組織の抽出液を作製し、これを培養したマクロファージと同様な機能を持つ樹状細胞に添加したところ、種々の炎症性サイトカイン(IL-23、IL-1β、TNFαなど)を産生するようになることが分かりました。脳抽出液をたんぱく質分解酵素処理すると活性能力がなくなることから、脳抽出液中の何らかのたんぱく質成分が樹状細胞を活性化させていることが分かりました。そこで脳抽出液を様々に分画して、最終的に質量分析計を用いた解析によってPrxファミリーたんぱく質が樹状細胞を活性化していることを突き止めました(文献3)。
Prxファミリーたんぱく質は、細菌から哺乳類まで幅広く存在し、通常は細胞内にあって過酸化水素を水に変換する酸化防止酵素として知られていました。したがって虚血などのストレスの際には大量に合成されて、細胞内の過酸化水素を水に変換して細胞を保護する働きがあると考えられていました。しかし細胞が壊死するとPrxは細胞の外に放出されます。私たちは虚血に陥った脳組織では細胞死に陥った組織の周りにPrxを含む細胞の残骸が集積しており、さらにマクロファージと接触していることを発見しました(図4)。これらのことは、虚血ストレスの際に細胞を生存させるために細胞内で大量合成されたPrxが発現されますが、いよいよ細胞死に陥るとPrxは細胞外に放出され、周囲に浸潤したマクロファージを活性化するというメカニズムがあるのではないかと考えさせます。
実際にPrxが樹状細胞を活性化するメカニズムを調べてみると、Prxは確かにTLR2とTLR4を介してマクロファージを活性化していることが分かりました。またPrxの活性を除去する中和抗体を脳梗塞モデルマウスに投与したところ、脳組織での炎症性サイトカインが抑えられ、梗塞体積が縮小し、神経症状が改善しました。
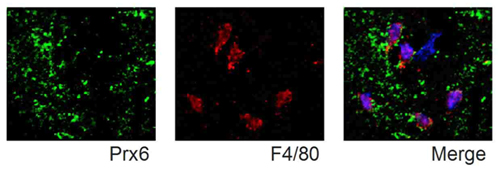
図4 Prxは細胞外に放出されてマクロファージを刺激する。
発症1日目の梗塞中心における免疫染色像を示す。左:Prx6を多く発現する多量のデブリス(壊死した組織の残骸)が存在する。中:F4/80陽性細胞(マクロファージ)が脳内に浸潤している。右:Prx6を含むデブリスがマクロファージの細胞表面上に共局在している。
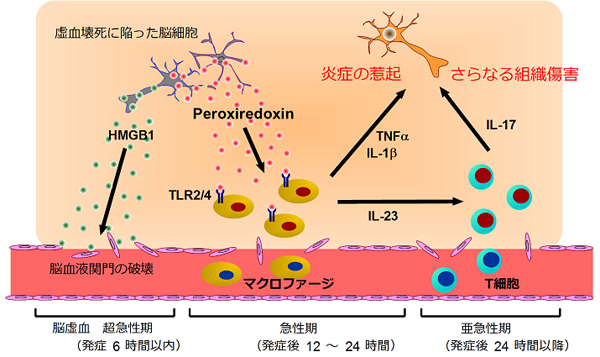
図5 脳虚血における炎症惹起因子としてのPrxの役割
脳虚血における炎症の経時変化を示す。発症6時間以内の超急性期には、虚血に陥った脳細胞からHMGB1が放出され、これは脳血液関門の破壊に寄与する。発症12~24時間以内の急性期にはマクロファージの浸潤が著明になるが、HMGB1の放出は見られなくなる。代わりに、虚血ストレスによって細胞内で発現したPrxが細胞死に伴って細胞外に放出され、浸潤したマクロファージをTLR2やTLR4依存的に活性化し、IL-23やIL-1β、TNFαを産生させる。IL-23はさらに遅れて脳内に浸潤したγδT細胞からIL-17を産生させ、さらなる組織傷害を惹き起こす。
今後の展開
“Time is brain”の言葉が示す通り、脳梗塞はできるだけ早期の治療開始が原則ですが、未だに脳梗塞の治療法は少なく、治療の開始が間に合わなかったり、適切な治療を開始していても神経症状が増悪したりすることがあります。そのために入院期間やリハビリ加療が長期化することになり、今後の高齢化社会に伴って医療経済を圧迫する可能性があります。本研究は、脳梗塞の新たな炎症メカニズムを明らかにしたものです(図5)。
今回の成果を踏まえると、脳虚血発症後の比較的早期に、γδT細胞やPrxを標的とした治療が開始できる可能性が示唆されます。さらに、炎症性サイトカインによる神経細胞の損傷のメカニズムについても理解が進むことが期待されます。
参考文献
1. Pivotal role of cerebral interleukin-17-producing gammadeltaT cells in the delayed phase of ischemic brain injury.
Shichita T, Sugiyama Y, Ooboshi H, Sugimori H, Nakagawa R, Takada I, Iwaki T, Okada Y, Iida M, Cua DJ, Iwakura Y, Yoshimura A.
Nat Med. 2009 Aug;15(8):946-50. doi: 10.1038/nm.1999. Epub 2009 Aug 2.
http://www.nature.com/nm/journal/v15/n8/abs/nm.1999.html![]()
2. Therapeutic effect of IL-12/23 and their signaling pathway blockade on brain ischemia model.
Konoeda F, Shichita T, Yoshida H, Sugiyama Y, Muto G, Hasegawa E, Morita R, Suzuki N, Yoshimura A.
Biochem Biophys Res Commun. 2010 Nov 19;402(3):500-6. doi: 10.1016/j.bbrc.2010.10.058. Epub 2010 Oct 20.
http://www.sciencedirect.com/science/journal/0006291X/402/3![]()
3. Peroxiredoxin family proteins are key initiators of post-ischemic inflammation in the brain.
Shichita T, Hasegawa E, Kimura A, Morita R, Sakaguchi R, Takada I, Sekiya T, Ooboshi H, Kitazono T, Yanagawa T, Ishii T, Takahashi H, Mori S, Nishibori M, Kuroda K, Akira S, Miyake K, Yoshimura A.
Nat Med. 2012 Jun;18(6):911-7. doi: 10.1038/nm.2749.
http://www.nature.com/nm/journal/v18/n6/abs/nm.2749.html![]()

七田 崇(助教)

左:七田 崇(助教)、右:吉村 昭彦(教授)
SHARE