慶應発サイエンス
生殖細胞のゲノムを守る小さなRNA分子
はじめに~生物の生死を左右する転移因子のはたらき~
私たちの体や細胞の設計図は、DNAと呼ばれる塩基配列の連なりから成り立っており、総じてゲノムと呼ばれています。ヒトゲノムは約30億塩基対という膨大な塩基配列情報を含み、46本の染色体と呼ばれる構造体に分散して格納されています(図1上)。細胞分裂を経てもゲノムの塩基配列情報は正確に複製され、それぞれの娘細胞に同じDNAが受け継がれています。ヒトゲノム解析の進展の結果、細胞を形作る上で必須なタンパク質は、約2万種類存在することが明らかとなっています。この2万種類、という数字も膨大なように感じますが、ヒトゲノム30億塩基対に対するタンパク質コード遺伝子の割合は、わずか2%に過ぎないことが分かっています(図1右)。では他の部分はどんな情報を持っているかというと、機能未知な領域も多いのですが、約45%は「転移因子」もしくは「トランスポゾン」と呼ばれる配列であることが分かってきています(図1右)。
次の疑問、ではなぜヒトゲノムでは転移因子の割合が非常に高いのでしょうか?それは転移因子の「転移」という性質に起因しています(図1左下)。転移因子は大きく2種類に分けられ、一つはDNAトランスポゾンと呼ばれるグループ、もう一つはレトロトランスポゾンと呼ばれるグループです。後者の転移様式はいわゆるコピー&ペースト方式であることが知られており、このコピーを作る能力によって、ヒトゲノムでは45%もの大きな領域が転移因子に占領されることになったと考えられます。このようなレトロトランスポゾンの転移は、ゲノムサイズの増加をもたらすことによって生物進化の原動力となったと考えられますが、その一方で、転移するゲノム内の位置によっては、生物を死に至らしめる原因ともなります。実際にトランスポゾンの転移が様々な疾患の原因となることが報告されており、例えば、がん患者の53%にトランスポゾンの転移が見つかっています。つまり、ヒトは転移因子の無秩序増殖の脅威と隣り合わせで生きている、と言えます。
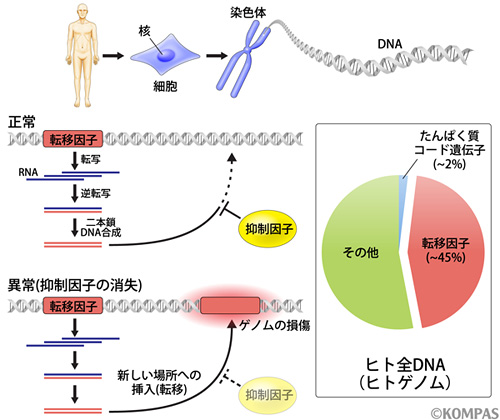
生命の遺伝情報は細胞内の核に存在する染色体に格納され、DNAという物質として書き込まれている(上)。ヒトゲノムは約30億塩基対から成り立つが、生命にとって重要な蛋白質コード遺伝子の割合は2%に対し、転移因子として分類されるものは45%もの領域を占める(右)。転移因子の中でも多くの割合を占めているレトロトランスポゾンはコピー&ペースト形式で自分自身の分身をゲノムの他の領域に挿入する能力(転移)を持つ(左下)。転移に必要な蛋白質は転移因子のメッセンジャーRNAがコードする。通常、転移を抑制する仕組みが働くことによって、ゲノムの品質が維持されるが、何かのきっかけによってこの抑制状態が消失すると、ゲノムの損傷が起こり疾患や細胞死につながる。
モデル動物を用いた転移因子の抑制機構研究
転移因子の多くは塩基変異によって転移能力を失っている場合が多いのですが、中には転移活性を持つものも存在しています。最近の研究によって、生物は転移因子を積極的に抑える仕組みを持っていることが分かってきました(図1左下)。本来生物が持つ転移因子の抑制機構を明らかにし、それをヒトに応用すれば、転移因子に起因する疾患を予防、もしくは治療することが可能になると期待されます。
私たちはモデル動物ショウジョウバエにおいて、2006年にpiRNAというRNA分子がPIWIと呼ばれる蛋白質と結合することを見出しました(文献1)(図2)。piRNAの特徴を解析したところ、転移因子の塩基配列と一致していたのですが、転移因子が数1000塩基から成るのに対し、piRNAは24から30塩基という短いものでした。また、転移因子がコードするメッセンジャーRNA(注1)とpiRNAは相補性(注2)を示したことから、piRNAはその相補性を利用して、PIWIを転移因子のメッセンジャーRNA上にリクルートするガイド分子として働くことが示唆されました。PIWIやpiRNAを人工的に欠損させると転移因子の発現が上昇し、生殖細胞形成不全が起こったことから、これらの因子は生殖細胞ゲノムを転移因子の脅威から防衛する役割を持つというモデルを提唱しました。このようにショウジョウバエにおいてpiRNAとPIWIという転移因子の抑制因子を見出したのですが、後に、ゼブラフィッシュやカエル、マウスにおいてもpiRNAが発見され、転移因子の抑制に働くことが報告されました(文献2)。しかし、マウスとショウジョウバエのpiRNA配列を比較すると、大きく異なっています。それはゲノムにおける転移因子の構成が生物種によって異なるなどに起因することがわかってきました。従って、個々の生物種におけるpiRNAの解析はその生物の転移因子の抑制機構を知る上で重要であると考えられます。
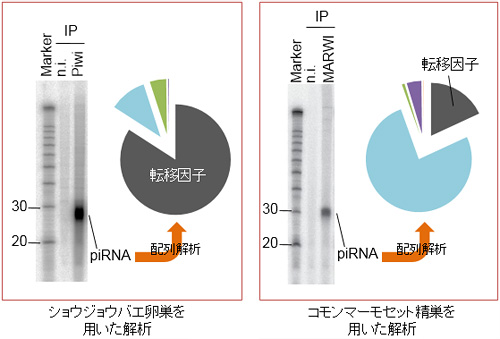
ショウジョウバエの卵巣とコモンマーモセットの精巣を単離し、その抽出液とPIWI蛋白質群を認識する抗体を用いて濃縮した。次に結合するRNA分子を可視化したところ、24-32塩基長のRNAが検出された(文献1及び3)。これを次世代シーケンサーによって解析した結果、転移因子に由来する配列を見つけた。ショウジョウバエと比較すると、コモンマーモセットの精巣では転移因子の割合が低いため、転移因子以外の遺伝子の発現を制御する可能性も示唆された。
コモンマーモセットを用いたpiRNAの探索
ヒトなどの霊長類においてpiRNAが存在することは予測されていたものの、詳細な機能解明はなされていませんでした。そこで今回私たちは、霊長類のモデル動物であるコモンマーモセットを用いてpiRNAの機能解明を、生理学教室の岡野栄之先生と実験動物中央研究所の佐々木えりか先生との共同研究で進めました。はじめにマーモセット精巣を単離し、その抽出液からPIWI蛋白質に結合する抗体を用いて、PIWI蛋白質(MARWIと名付けました)とそれに結合するRNA分子を精製し、piRNAと結合しているかどうかを検討しました(図2)。その結果、MARWIはpiRNAと結合していることを見出しました(文献3)。次に、piRNA配列を次世代シーケンサー(注3)と呼ばれる膨大な配列数の解析が可能な装置を用いて決定しました。その結果、MARWIに結合するpiRNAの中に、転移因子由来の配列が存在することを見つけました。従って、霊長類においても他の生物種同様にpiRNAによる転移因子の抑制機構が備わっている、と考えられます(図3)。
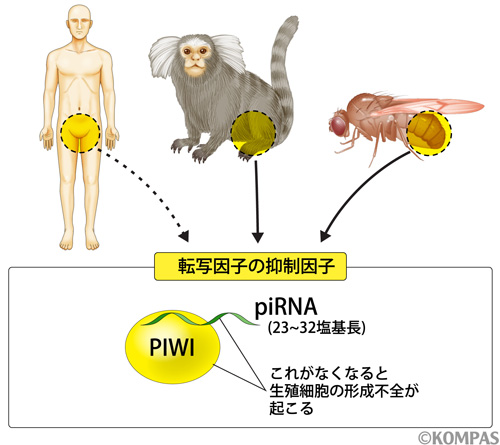
piRNAはPIWIと呼ばれる蛋白質と結合するRNA分子であり、これが消失すると転移因子の発現が上昇することがショウジョウバエやマウスで報告されている。今回の解析によって、霊長類モデル動物であるマーモセットにおいても転移因子由来のpiRNAが見つかった。従って、piRNAは様々な生物種に共通の転移因子の抑制分子であると考えられる。現在、piRNAの多くは生殖細胞で見出され、生殖細胞の形成維持に必須であることが分かっており、ヒト不妊症との関連性がないか精力的に研究が進められている。
今後の展開~RNA分子を介した疾患予防法の開発~
我々の研究から、ショウジョウバエから霊長類まで様々な生物種において、転移因子由来のpiRNAが発現していることが明らかとなりました。従って、RNA分子を介したゲノム防衛システムは種を越えて保存されたメカニズムであると考えられます。生殖細胞は次世代にそのゲノム情報を受け継ぐ重要な細胞です。そのような細胞では、特に転移因子の抑制が重要になっている、と考えられます。今後、piRNAの作動メカニズムや合成メカニズムが明らかとなれば、人為的にpiRNAを作り出すことで転移因子を抑制し、疾患を予防することが可能になると期待されます。
【用語解説】
注1)メッセンジャーRNA
蛋白質のアミノ酸配列を規定する塩基配列を持つRNA分子。DNAを鋳型として合成(コピー)される。蛋白質はメッセンジャーRNAのコドンと呼ばれる3塩基の並び配列に従って合成される。従って、メッセンジャーRNAは遺伝情報を伝える重要な中間物質として働く。
注2) 相補性
DNAやRNAの塩基対合の安定度。DNAの場合、アデニンとチミン、グアニンとシトシンがそれぞれ塩基対を形成する。一方RNAでは、アデニンとウリジン、グアニンとシトシン、そしてウリジンとグアニンが塩基対を形成する。塩基対の長さが長いほど、その二本鎖は熱力学的に安定となる。piRNAの長さは25塩基程度であり、これは完全に相補的なRNAと37℃において安定な二本鎖を形成し得る長さである。
注3) 次世代シーケンサー
数億から数千億塩基の膨大な塩基配列情報を一度に取得できる装置。従来型のシーケンサーでは約100種類程度のDNA断片を1台の装置で同時に解析することが限界であった。それに対して次世代シーケンサーでは、数千万以上のDNA断片を並列して処理する技術の開発がなされ、大量の塩基配列情報を得ることが可能となっている。2003年にヒトゲノムの解析がほぼ完了したが、これは従来型のシーケンサーによって為され、ほぼ10年を要した。しかし、次世代型シーケンサーを用いれば、1週間以内でヒトゲノムを解明できるようになっている。この技術は様々な分野で応用され、生命現象へのアプローチを大きく変化させている。
参考文献
- Specific association of Piwi with rasiRNAs derived from retrotransposon and heterochromatic regions in the Drosophila genome.
Saito K, Nishida KM, Mori T, Kawamura Y, Miyoshi K, Nagami T, Siomi H, Siomi MC.
Genes Dev. 2006 Aug 15;20(16):2214-22.
http://genesdev.cshlp.org/content/20/16/2214.long
- PIWI-Interacting RNA: Its Biogenesis and Functions.
Iwasaki YW, Siomi MC, Siomi H.
Annu Rev Biochem. 2015;84:405-33.
http://www.annualreviews.org/doi/abs/10.1146/annurev-biochem-060614-034258 - Small RNA profiling and characterization of piRNA clusters in the adult testes of the common marmoset, a model primate.
Hirano T, Iwasaki YW, Lin ZY, Imamura M, Seki NM, Sasaki E, Saito K, Okano H, Siomi MC, Siomi H.
RNA. 2014 Aug;20(8):1223-37.
http://rnajournal.cshlp.org/content/20/8/1223.long

SHARE